a) What is the distribution and abundance of the prey of larval fish within the Bay and how does this vary within the Bay as the salinity and temperature distributions change throughout the seasonal cycle?
Grazing rate is measured by the gut fluorescence method. The ingestion of phytoplankton by each copepod taxon will be determined from the product of ingestion rate per copepod and the abundance of that copepod. Grazing by other numerically abundant mesozooplankton is estimated by applying either literature values for grazing at similar temperatures or size specific rates estimated from the copepods to the abundances derived from our net tows.
Community grazing rates determined at specific sites will be scaled up to larger areas based on more extensive measurements of zooplankton abundance and concomitant water properties and the degree to which taxonomic distribution and abundance can be determined by hydrographic variation determined using multivariate statistics like correspondence analysis.
Larval fish are sorted from 150um neuston tows to obtain individuals for gut contents analysis. Individuals are measured (notochord length) and their guts excised, measured (width) and identified. The guts of only morphologically intact individuals will be included in the analysis. Identifiable gut contents are be compared to the density of food organisms enumerated in net tows, sieve samples and whole water samples. The same procedures are being used with small-mouthed juvenile and adult fishes captured in Fla. DEP trawl and seine net samples.
Using Acartia tonsa collected in Biscayne Bay, we amplified a partial region of the large subunit of ribosomal DNA using the PCR (polymerase chain reaction) method. We then sequenced this region on a LiCor automated sequencer and isolated a unique species-specific area of the subunit. A non-isotopic DNA probe was then prepared for that oligonucleotide sequence signature. This probe is used to identify unrecognizable larval fish gut content fragments with a competitive PCR detection technique. The approach is not only qualitative but quantifiable.
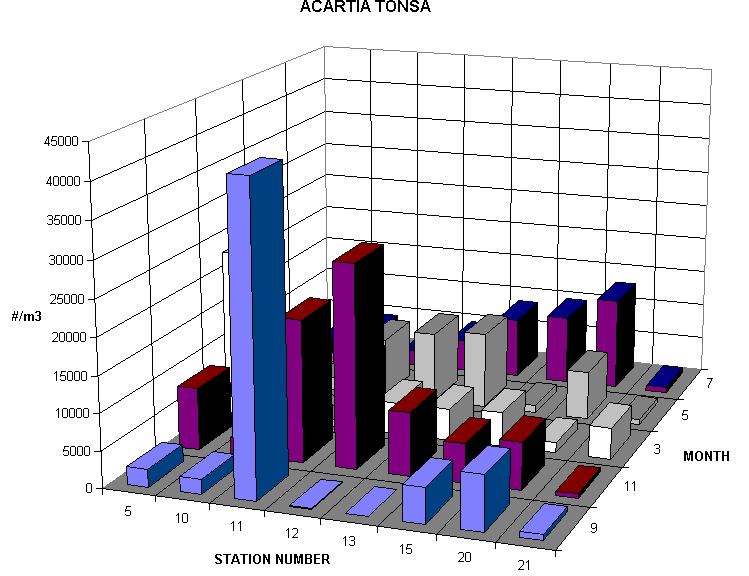
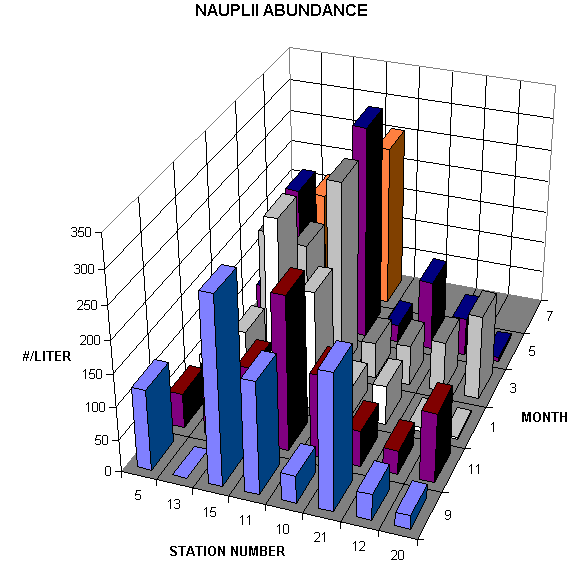
The most abundant net caught copepods were Acartia tonsa, Oithona nana and Paracalanus crassirostris. In September when salinity ranged from 33-43ppt Acartia abundances ranged from 16 to 8216/m3, Oithona from 307 to 40460/m3, Paracalanus from 10 to 785/m3. In November when salinity ranged from 22-32 ppt. Acartia abundances ranged from 79 to 5948/m3, Oithona from 427 to 29952/m3, Paracalanus from 204 to 15384/m3. Only Paracalanus was significantly more abundant in November. In the one January station already analyzed salinities were slightly lower than in November and Acartia abundances were considerably higher (fourfold or more in comparable 150um net tows). In September, bucket sampled copepod nauplii ranged from 21 to 284 liter-1 while in November from 36 to 235 liter-1. The average length of a copepod nauplius was 116 um and the average width was 55 um. Such small nauplii are expected since the dominant copepods are comparatively small but from morphological examination alone it is difficult to determine the species of the nauplii enumerated. Other copepods that were common but much less abundant included Tortanus setulosis, Euterpina acutifrons, Longipedia helgolandicus and Calanopia americana.
The most abundant meroplankton in net samples were gastropod larvae and pelecypod larvae. While more highly variable than the copepods these were on occasion the most abundant organisms in the 64um net tows. In September they ranged from 553 to 69,732/m3 and in November from 42 to 187,436/m3. Other meroplankton included zoea, decapod larvae, echinopluteus and heteropod larvae.
Comparison of 150um mesh tows taken during the day and at night indicated that large Acartia (0.5-1.0mm body length) and Calanopia americana were significantly more abundant. In fact Calanopia was rarely sampled during the day. The abundances of Oithona, Paracalanus, Longipedia and Euterpina and chaetognaths were insignificantly different night and day. In other shallow estuarine systems older Acartia are reported to have a semi-demersal behavior, staying close to or on the bottom during the day and rising up into the water column at night. Because Acartia is the largest of the dominant species and is well known to have a high tolerance for varying salinity, additional night-day comparisons are planned.
{kind=link}
{kind=link}